
Readers of this series may have detected a slight yet pervasive diminution of the import of the famed "killing claw" over the course of these posts. In my first post I documented the shift in scientific thought on these claws from scythes that cut meter long slashes in prey to crampons that allowed hitching rides on the hides of dinosaurs to ultimately the prevalent modern interpretation of raptor prey restraint (RPR) model of Fowler et al. in which prey subequal in size is grasped by all four digits. I reiterated a point seldom mentioned from the Fowler et al. paper on the dromaeosaurid RPR hypothesis: relative to accipterids, the ungual grasping ability of dromaeosaurids was >not as strong< as these birds in that arena i.e. they were not simply scaled up hawks. Later in that post I suggested a role for the arm/wings for pummeling prey/combatants as the feet grasped and pinned the animal. In my next post focusing on aggressive/combative scavenging in these animals I focused in on digit II as a useful tool in pinning large carcasses down as the head, neck and teeth pulled back on flesh - an idea supported by the unique morphology of the denticles on these theropods and the presence of enlarged digit II claws in several birds that work in a similar fashion.
My contention is that the import of digit II - so highly regarded that it is referred to as the "killing claw" - has both culturally and scientifically influenced these animals to the point that other aspects have been enshrouded. But was the "killing claw" really the most pivotal aspect of these animals behavior and ecology? I think not, or at least >not always<.
If the use of the "killing claw" digit II was indeed the be all and end all of dromaeosaurid prey capture and feeding technique we should be able to make some predictions to test that assertion. That over the course of the 100 million year evolutionary trajectory of these animals an increasing reliance on ungual prey capture will 1) show a trend towards shorter and therefore stronger legs i.e. less cursorial adaptations 2) as firepower is concentrated in the feet for killing the robustness of the skull and teeth should hold steady or potentially diminish. In the early Cretaceous Deinonychus we have a relatively sub-cursorial but highly adept foot grasper - again there is a bit of an inverse relationship between foot grasping strength and cursorial ability as I discussed in the first post and which Fowler et al. highlighted in their paper. I will cut and paste the source of this observation from the Fowler et al. paper:

As evidence for the purported trend in increasing foot strength Fowler et al. cite Deinonychus (early Cretaceous), and Velociraptor & Saurornitholestes (late Cretaceous). While Deinonychus and Velciraptor have relatively short metatarsi I can't see how they interpret the leggy Saurornitholestes as an example of this trend. Additionally there are some notable omissions, most obviously the name sake for the whole family Dromaeosaurus!!
In Dromaeosaurus albertensis cursorial adaptations are highlighted, the killing claw is relatively atrophied, and the skull is relatively massive and robust (almost tyrannosaurid like as GSP has commented). I mean just check out the skull of this animal, there is nothing slight, superficial, or atrophied about it at all:
 |
| robust head of Dromaeosaurus albertensis. credit LadyofHats. public domain |
 |
| relatively diminished claw size/strength D. albertensis. credit LadyofHats. public domain |
Several of these trends towards diminished ungual strength and/or increasing skull robustness also play out in Dakotaraptor (cursorial w/diminished foot grasping ability) and the very robust skulled Atrociraptor.
 |
| Atrociraptor credit Ferahgo the assassin (Emily Willoughby) CC3.0 |
No dromaeosaurids were not evolving protobeaks or going edentulous despite the persistent artistic meme and no they were not diminishing emphasis on teeth and jaws.
It really is all about the teeth....
To drive home this contention I want to revisit a famed piece of data that has caused quite a stir in terms of whom and how it was done - the famed Tenontosaurus bite marks and the case for Deinonychus "bite strength". A technical paper by Gignac et al. (2010), a blog post by Mark Witton, a blog post by central coast paleontologist, and an internet article/summary from The World of Animals all highlight the attention and thought these remains have attracted.
That these bite marks have evolved into a bit of a paleontological "who done it" has always irked me. Not because of a lack of data or some systemic problem with the analysis - but because of the pervasive "explaining away" of data that most parsimoniously points to Deinonychus as the perpetrator. Several ideas have been bandied about in an attempt to account for these bite marks by Deinonychus, a predator that appears to not have an especially high bite force.
Let's unpack them:
1) An undescribed and undiscovered tyrannosauroid dinosaur did this damage.
We have seen this story before... tremendous damage to bone - no way a "blade toothed" theropod did it much less a puny dromie. Let's just imagine a stout toothed, bone crunching tyrannosauroid existed at a time when such animals were basically all blade toothed anyways, and make this essentially fictional animal the perpetrator. Made up tyrant lizards did it!! Pesky blade toothed theropods just stay in your lane - you guys can't bite through bone the way tyrant lizards can!!
As you can tell (snark alert) I am not so much a fan of this idea. We have evidence of Deinonychus being the most ubiquitous theropod in the area; the tooth arcade matches; broken teeth in the area; the well established Tentontosaurus - Deinonychus relationship - the whole tide of evidence points to Deinonychus. If a cryptic lineage of stout toothed, bony crunching tyrannosauroids existed at this time I will be happy to be proven wrong - as of now I know such evidence and of the tyrannosauroids from this time period they are blade toothed predators without expanded jaw musculature - although I have heard murmuring of tyrannosauroid teeth from the same formation (but blade toothed not lethal bananas).
2) Deinonychus could bite hard, but it did so extremely rarely.
I mean really? Remember when you kept hearing how humans only use 10% of their brain? Yeah, this explanation sounds a lot like that. Over designed with a bite force exceeding modern American alligators yet barely ever uses this strength? I can't really go with this thought.
3) Stronger bite than predicted from studies.
I don't think that this animal had much of a stronger bite than studies indicate. I believe that we have been a little bit more than led astray by always looking at static bite strength as opposed to other methods of cutting that highlight speed, friction, and getting those darn denticles to do the work for you. It really is all about the teeth and it really is all about getting the denticles to work in a way that maximizes cutting efficiency with minimal effort and wear & tear of the tooth.
Its high time we start looking at hypotheses that invoke Deinonychus as the prime perpetrator. I will put out a hypothesis that highlights an unorthodox feeding mechanism in these animals, that is consistent with the data, and offers much explanatory power for the observed data.
To prime you for it I want to look at birds a bit (as usual). To really confound the situation the obvious choice is flamingoes - because what better to compare dromaeosaurids to than flamingoes, amirite?!?
I mean, excuse the poor video quality, but just look at those tongues go!! It is the tongue just pumping back and forth causing the whole neck to just vibrate. I have no idea why these flamingoes engage in this lingual vibration? Anyone ever see wild flamingoes do this? I would have to assume that they pump their tongues back and forth to filter food but in my observations of these captive Chilean flamingoes they just do it while walking around... probably just bored.
No I am not suggesting that Deinonychus had some sort of lingual vibrational apparatus set up - just pointing out how one muscular organ - the tongue - can move with such speed and power in this bird that it vibrates the whole head and neck of these animals. I mean can your tongue move with such speed and power that it causes the whole body to hummm and vibrate... ummm never mind. The message I am trying to convey here is that when we look at avian feeding mechanics - and by extension many dinosaurs and especially paravaians/maniraptorans/dromaeosaurids - there is a lot of potential for quick twitch muscle, full body and/or neck movement involved in the feding apparatus. To drive home this point, literally, what would woodpeckers be without their exceptionally quick and rapid - fire neck movements? Yes, it is the skull of woodpeckers that is wonderfully equipped to handle the blows and stresses incurred but without the power and speed provided by the neck the woodpecker would, essentially, not peck. It would just be a bird with a strong skull.
An often overlooked aspect of feeding mechanics is elaborating on how parts of or the whole of the body is engaged in feeding mechanics - the head need not be looked at as an isolated aspect of the process. Regular readers should note that I have made this point before on antediluvian salad especially with regards to twist or torsional feeding (death rolls) in plesiosaurs and in my bonesaw shimmy hypothesis on Allosaurus in which it is rapid neck movement in both the fore and aft direction that allows the denticles on the front and back end of the tooth to saw right through tissue. Bite force was not especially important in that hypothesis, in fact tight clamping would work against free movement of the denticles over the tissue.
This hypothesis does take some inspiration from the bonesaw shimmy model but it does deviate from it in several ways.
I propose that fast twitch muscular contractions of the neck, torso, and even tail would pulse out vibrational waves of energy towards the head. As bipeds that do not have their front feet on the ground these pulses of vibrational energy would travel unhindered through the neck, head, teeth, and ultimately into the food item they are cutting into. As the vibrational energy literally vibrated the tooth back and forth into the food item the peculiar denticle pattern of dromaeosaurids comes into fruition as an optimized adaptation to literally bore and auger into tissue.
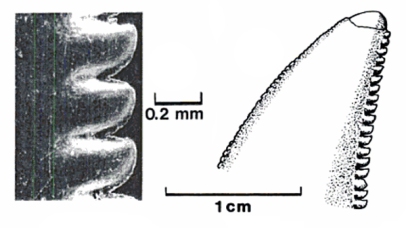
The most striking and unique feature about the denticles on Deinonychus is that they are fairly reduced on the front of the tooth but very pronounced on the rear. But even stranger is the manner in which they are curved on the rear side which is towards the tip of the tooth, referred to as apical hooking. Fowler et al. suggested that this unique denticle design would optimize cutting into tissue as the prey animal was held in the RPR model and the head of the dromaeosaurs was sub-vertical with the nose facing down and biting between the legs. However this suggestion by Fowler fails to address the issue that many other theropods likely held prey/food down with their feet and wrenched off bites in a sub-vertical manner yet these theropods did not evolve such weird denticles as seen in many dromaeosaurids.
But if we imagine each denticle as a "tooth" and each tooth having a respective duty in food processing a potentially new perspective emerges that could explain the unique bone damage ascribed to Deinonychus.

As the piece of food is grasped a strong bite is first established. The slight and reduced serrations on the front of the tooth are useful here in establishing a piercing bite - not very deep as their bite force was modest but merely a small indentation into the article of food. Once a purchase is made then the body commences vibrations - potentially a combination of head, neck, torso, and tail rapid fire twitches - which allow the tooth to bore and auger into the food particle i.e. bone. As the "bore hole" phase commences the utility of the weird apically hooked denticles comes into play as each denticle literally chips and shreds away at tissue like individual teeth. As the tooth works its way into the material it leaves a remarkably accurate impression of the tooth - a literal bore hole that for all intents and purpose can be read as a puncture. Once the integrity of the material is weakened substantially the item can be pinned with the arms and/or feet and the head and neck are pulled back strongly incurring further and more drastic damage as the tooth is dragged back through the (weakened) material literally leaving deep bone raking marks and furrows. It is also potentially possible that vibrations of the body were not emphasized or were in fact used in concert with multiple quick bites - essentially chattering of the jaw - in which micro - abrasions from the denticles work to carve into tissue.
This "vibrational feeding" hypothesis could potentially explain the two types of feeding traces recorded on the bones of Tentontosaurus which include longer gouges and simple punctures.

Above you see the type of "bone rakings" I mentioned earlier. An initial puncture is established and with the teeth embedded now the neck and body can pull back and rake through tissue.
What I suggest was occurring here is that these were investigative bites into bone. The theropods were gouging into the bones to see if there was ample nutritional value in them to justify the effort and potential wear of teeth. There would always be a three-way tradeoff between nutritional value versus the effort and wear on the animals feeding apparatus all of which is tempered by the relative health of the animal i.e. how desperate for food is it. Ultimately it looks like the theropods abandoned the bone consumption in this case.


That dromaeosaurid teeth occasionally show extreme wear - especially on the tips as should be predicted in this model - is very interesting.
 |
| worn tooth "Dromaoesauroides" wiki |
 |
| Private "dromaeosaur" tooth Montana .84" |
 |
| Judith River "dromaeosaur" tooth |
Of course it is worth mentioning that there is a lot of room for deviation in this model and we need not assume that all dromies employed vibrational feeding to the same extent. Indeed Dromaeosaurus could have employed a lot more emphasis on traditional "power chomps" than what I suggested for Deinonychus.
In theropods, being both ziphodont toothed and bipedal, there is no go to analogy among modern tetrapods - birds don't quite tell the whole story and neither do monitor lizards. So maybe we should expect some unothodox feeding mechanics.
Lifestyles of not only the quick and cursorial but the slow and persistent as well...
And now for the tail. Probably the aspect >least likely< to be assumed to be involved in "making dromaeosaurids nasty again". But it is the tail that is the most important aspect of these animals I will argue. The tail is what really pulls together all the disparate attributes of these animals and makes them what they were. And what they were was quite literally the most successful in tenure small to medium sized terrestrial hunter - scavengers that have ever existed. A unique blend; of accipterid "raptor"; combative scavenging vulture; bone chomping hyena ; a dash of felid; and, yes, highly efficient cursors similar to kangaroos, hyenas, humans, wolverines, and Arctodus.
One of the persistent ideas that has gained popular recognition in recent years is that dromaeosaurids were sub-cursorial - that they were slow. A chief argument put forth to support this notion is that the ankle bones - the metarsii - were rather short. And this is true for many species - Deinonychus and Velociraptor in particular - that were gaining mechanical advantage of foot claw strength at the expense of speed. But this was not so true in several other species - Dakotaraptor and Dromaeosaurus for instance - that were leggy to an exceptional degree. I am just not at ease with suggestions that species at the lower end of the spectrum were heavy footed clunkers - they could probably all put on a decent burst of speed if need be. Ursids (da' bears) have all the hallmarks of real clunkers but put on good speed with their short ankles. Keep in mind that dromies were competing with larger - and in the case of tyrannosaurids likely larger and quicker - theropods as well as azhdarchids. It is not always about being the fastest - but about being more agile when fleeing a larger threat. With their arm - wings and long tails doubtless many dromies frustrated an angry tyrannosaurid back in the day with their superior agility.
The dromie tail, just like the dromie "killing claw" has gone through a twisted and convoluted history of interpretations and revisions. A brief recap. Ostrom interpreted the tail as an intricate balancing rod that facilitated use of the "killing claw" for kicking and hanging onto prey. Each subsequent interpretation of dromie killing technique from hanging onto the side of prey and biting to the RPR method invoked the tail as intricate balancing organ for their respective prime foraging technique.
To add further context to the strange saga of dromie tails I want to revisit a post from Pterosaur.net Blog (remember that great site?) Dragon Tails: What Pterosaurs Teach Us About Velociraptor that made the strange and startling comparison between dromaeosaurid tails and rhamphorynchid tails... wtf? Well there is a comparison to be made there and it is not soooo strange when we work from the starting point that dromaeosaurids likely had flighted ancestors... so that they inherited a tail that - presumably - shared a convergence in form and function with rhamphorynchid pterosaurs.
 |
| credit Scott Hartman used w/permission . blog Scott Hartman's Skeletal Drawing |
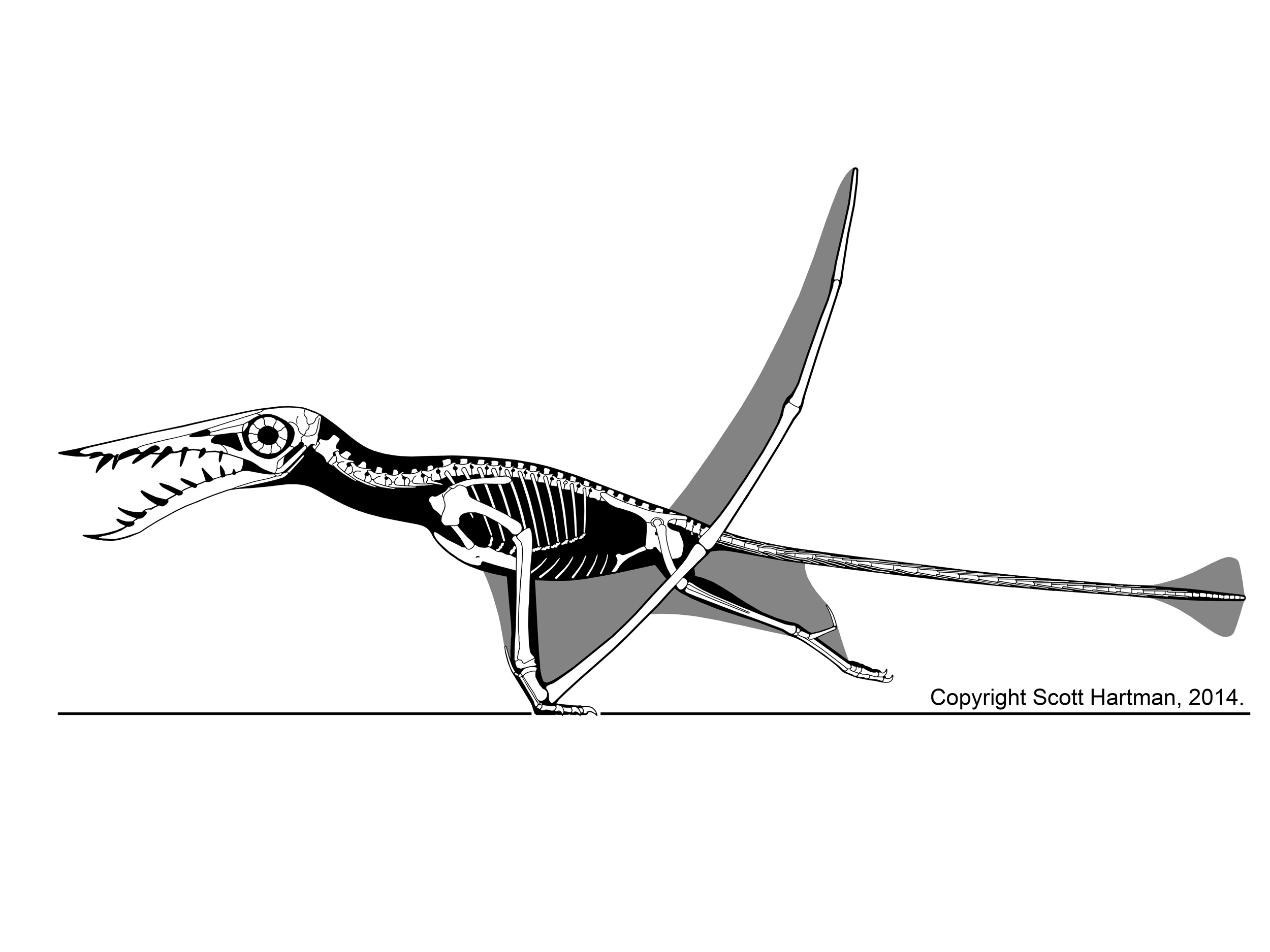 |
| credit Scott Hartman used w/permission . blog Scott Hartman's Skeletal Drawing |
So if dromaeosaurids inherited the weird morphology of their tails from flighted ancestors - full of chevrons, diminished musculature, partially ossified dual tendons (i.e. caudal rods) there becomes two rather interesting questions: 1) what adaptive benefit did these features incur in flighted dromaeosaurids and tailed pterosaurs? and 2) how was this morphology coopted into terrestrial based dromaeosaurids? Question number #1 I am going to leave alone but I think it is a long overdue question that needs analysis but question number #2 is what I am going to approach here.
 |
| credit Scott Persons |
What I am going to suggest is that dromaeosaurids across all ranges of absolute speed and leg length - were highly efficient long distance pacers. They could and did just keep going for miles at a time at a relatively moderate pace. The whole lot of 'em could just run you to death. And the key to this long distance efficiency was the tail. The tail - the whole organ - served as an elastic recoil that allowed these animals to store, redistribute, and recoup energy for efficient, long distance traveling. I have seen scant attention to the tail as an aide in terrestrial efficiency in dromies. Despite the fact that these animals were terrestrial and the tail of dinosaurs is intimately linked with movement - especially per the caudemofemoralis muscle. Darren Naish raised the question of dromie tails back in 2008 (What the hell is going on with dromaeosaur tails?) in light of Norell & Makovicky (1999) describing an articulated and sinuous Velociraptor tail. The comment section is interesting. I do note in it a pervasive sentiment of trying to "explain away" the sinuous tail - the presupposition being that stiff tails is the better supported null in dromies to start with. But is a stiff tail the better supported null or is it just how we grew up expecting dromie tails to behave? In either case lateral flexibility shown in both Velociraptor and Bambiraptor seems to have prevailed. But there is one comment by Alan #19 that I believe was very prescient and which received literally no attention in the discussion.

I think Alan was on the right track as goes energy efficiency although I doubt the hopping dromie scenario has much merit - indeed trackways have proven otherwise.
From my own experimental paleontology in which I strapped on a huge tail to my butt at SVP Los Angeles and commenced to simultaneously entertain and annoy attendees I noted several patterns. What was really interesting to me is how much that darned tail moved around. Literally the smallest movement I made would thoroughly send the tail in motion. And what was most notable was the dramatic up and down oscillations that the tail went through as I walked. Each foot fall would create a simultaneous rise and fall of the tail - even the smallest and daintiest step. Don't believe me strap one on yourself and be a dinosaur for a day - you'll see what I am talking about.

These up and down movements of the tail that occurred simultaneously with each footfall likely occurred in all dinosaurs to some degree. What is interesting is that dromaeosaurid tails - because of their "caudal rods" - were designed to diminish this up and down movement of the tail as thoroughly explained by Scott Persons on his post on dromie/rhamphorynchus tail convergence. Note in the pic below how the caudal rods are neatly stacked against one another on the vertical plane to limit movement dorso - ventrally.
 |
| Caudal rods in Deinonychus prevent up and down movement of tail credit Scott Persons |
 |
| Bambiraptor tail credit Scott Persons |
This model of locomotory efficiency is not without parallel in animals that have to move across vast expanses to find and locate rare and ephemeral food resources. A leading hypothesis concerning Arctodus is that it was a highly efficient long distance pacer that scavenged and usurped carcasses (Matheus, 2003) utilizing long legs and elastic recoil to travel at a moderate pace over long distances. Hyenas have long been noted for their efficient loping pace that allows large scale movements and carcass retrieval. Kangaroos and wallabies are well noted for their ability to travel long distance at an extremely energy efficient pace owing a lot to the elastic recoil in their leg tendons. Indeed a robotic kangaroo has been designed that utilizes such elastic recoil in the tail to recoup energy for movement.
I would be remiss not to mention the endurance running hypothesis has been invoked as a strategy for both scavenging and pursuit hunting in our own genus aided by the achilles tendon. To further quell the notion that short legs - such as in Deinonychus or Velociraptor - imply a suboptimal terrestrial movement capability let us not forget about wolverines which are notorious long range hunter -scavengers despite being very short limbed. I don't know if there have been any studies on the locomotory efficiency of these animals but I suspect there is something to 'em in those regards. There are at least loads of references to the marathon travels of these facultative scavengers.
"It is absolutely impossible for any human to keep up with a wolverine. What wolverine can do is just beyond human."
"A wolverine crosses a topo maps like we cross a street."
"They devour the landscape at a constant 4 mph regardless of terrain."
An energetic bundle of tooth, claw, and attitude? Switching from small game foraging to large carcass acquisition as the seasons dictate? Bone consumption? Able to outpace, outcompete, and outwork competitors that are several orders of magnitude larger in size? Thriving in areas and desolate habitats that other predators eschew (snowfields analogous to dune fields in these regards)? A little bit of the Gulo gulo in your dromie? You bet.
Making dromaeosaurids nasty again... Invoking the wolverine as a likely analogue for many dromies, it doesn't get much nastier than the demon of the north.
Ichnology: What Does It Tell Us?
Xing et. al. (2013) document a variety of dromaeosaurid trackways from the lower Cretaceouls Hekou group in China. The pace was not very high at about .75 meters/second which is about 1.7 mph or 2.7 km/hour. Average human walking speed is said to be about 3.1 mph or 5.0 km/hour. Let me just cut and paste the discussion:

So although these particular dromies seem to be moving along at slowish pace - perhaps they had full bellies or were just walking down for a drink. It is noteworthy that they mention several dromie ichno-species in the last paragraph that seem to be cruising along at quite brisk paces and one zipping along pretty good.
Dromaeosauripus from Korea at 4.86 m/s (Kim et al. 2008) which is 10.9 mph / 17.5 kmh
Paravipus (Murdoch et al. 2010) at 1.67 m/s and 3.61 m/s which is 3.6 mph / 5.8 kmh and 8.1 mph / 13.0 kmh
Dromaeopodus at 1.63 m/s (Li et al. 2007; Kim et al. 2008) which is 3.6 mph / 5.8 kmh
Considering that to document an actual predatory chase in the footprint record is exceptionally rare and that there is no evidence that a chase was in progress in any of these instances the ichnological data is very interesting. We see a range of speeds here from the more leisurely .75 m/s to a quite hectic 4.86 m/s. If we assume that these are reasonable cruising speeds then the small sample size we have does point to a relatively fast paced "cruiser" similar to humans, wolverines, coyotes, and hyenas that can cover vaste expanses of land at an efficient pace as the penultimate terrestrial hunter - scavengers of their time.
The Seldom Mentioned Fact of Dromie Toe & Heel Pads
The trackways from this study demonstrate that dromies had big ol' foot pads like two toed ostriches but also large heel pads! So pay attention to this aspect paleo-artists >at least some< dromies had big fat derpy looking foot/heel pads that are universally never depicted at all or large enough in paleo-art depictions (including my own). Why has this well documented aspect of dromaeosaur foot anatomy never penetrated into popular depictions? I mean no one - literally nobody - including world renowned paleoartists or more obscure/enthusiast artists depicts dromies with large heel pads. Yup the toes had big padding but the heel pad would have been very apparent in life. And this is from a peer reviewed paper with several notable authors including most notably to my western biased eyeballs, Phil Currie (who is btw the last author).

Such fleshy and large toe/heel pads would assist in stalking behavior by muffling sounds, stability, absorb stress from cursorial activity but I also have to wonder if such fleshy structures would diminish grasping effectiveness?

Also check out the base of digit II often reveals a bit of a fleshy toe pad. Dromaeosauripus yongjingensis represents a fairly large "Utahraptor" size dromie but other dromie footprints reveal fleshy toe pads and heel pads.

Kim et al. did a paper with reference to a speedy little dromie in the above discussion (Kim et al. 2008) from Korea of Dromaeosauripus moving along at about 4.86 meters per second (10.9 mph or 17.5 kmph).
Abstract:

I also should give some space to the ichnological data pointing to at least six large dromies traveling in parallel and the special emphasis the authors give to the toe and heel pads in the footprints ( Li, 2007).



So when depicting the average large terrestrial dromie foot think more about ostrich feet than harpy eagle feet. Except that unlike ostriches dromies often had big ol' heel pads in addition to toe pads that would have further cushioned the foot and added a degree of stability normally not ascribed to these animals. The increased surface area would have facilitated greater efficacy and stability of movement in dubious terrain such as dune fields and mud flats.
 |
| ostrich foot credit Masteraah CC 2.0 |
A lot of questions to be answered but I do think that a fresh appraisal of these animals as primarily terrestrial long distance hunter - scavengers that have to cover a lot of ground efficiently is needed. Optimal walking versus optimal cruising speed can be addressed with larger sample size of ichnological data and computational methods... What I can say is that the anatomy of the tail likely has something to do with terrestrial locomotion and efficiency of gait is as good of a hypothesis to investigate as any...
These animals had to have been able to move and move well. They had to have traversed wide distances to secure meals in often times inhospitable terrain. They had to have competed against larger and aggressively hungry and growing youngsters of tyrannosauroids, carcharodontosaurids, and other theropods. They had to get to carcasses before large pterosaurs got all the good stuff. They had to have been at least reasonably competent in these realms to have persisted as... I don't know... the longest tenured group of small - medium sized tetrapod terrestrial hunter - scavengers that ever existed ( I know I said it before but it bears repeating). Speedy thieves indeed.
Earlier in this article I suggested that there was a link between the tail and biting apparatus in these animals - that their functions dovetail together. At the risk of piling one hypothesis on top of another let me put it out there that the diminished dorso-ventral movement of the tail as dictated by the caudal rods would have shunted more of the potential energy towards the anterior of the body - essentially towards the head, jaws, and teeth - during vibrational feeding.
 |
| credit Duane Nash Tsaagan & Velociraptor |
Final Thoughts
Both a scientific and cultural emphasis on the "killing claw" in dromaesaurids has obscured a more nuanced, multifaceted, and holistic approach to these animals; that the "ground hawk" model has so embedded itself into our conscious; that the potential role of arm-wings as brutal spiked clobbering devices analogous to wing pummeling in modern aves has been overlooked; that the teeth were highly specialized and brutal weapons in their own right capable of extreme insults to carcass integrity (including bones) and perhaps full body "vibrational feeding"; that the importance of head and tooth weaponry did not diminish over the evolutionary history of this group but sometimes increased while emphasis on "killing claw" and foot grasping capability did in fact sometimes diminish; that cursorial ability did often times increase in capability and that all dromaeosaurids may have benefited from elastic rebound provided by caudal rods in the tail enhancing long distance, mid-paced terrestrial efficiency of movement as well as large fleshy toe & heel pads; that life appearance may have been more varied than simply "grounded hawks" with "dapper" haircuts but imbued with much of the panoply of life appearance we see in ratites, predatory and scavenging accipterids, cathartidae, bucerotidae, galliformes, and other large/terrestrial aves including but not limited to large exposed fleshy areas including caruncles, wattles, frills, dewlaps, and other tough - elastic - and fleshy skin derived outgrowths for thermorgulation and sexo-social signaling; that these attributes when generously applied to an outstanding and long lasted dynasty - in fact the longest tenure of small to medium sized tetrapod terrestrial hunter - scavengers to have ever existed - create a strikingly original, efficient and for lack of a better term "nasty" eco-morphological package that punched above their own weights in many categories.
They were above all else... awesome... bro.
And finally... can we please stop calling them raptors? That name is already taken!! You may have noticed through the course of these articles that I have bounced a lot between dromaeosaurid and dromie... I probably in retrospect should have used the term eudromaeosaurid through out as they are what I am principally talking about here not microraptorines or unenlagines.
I vote for calling these guys "dromies"and am fully favor of eschewing the befuddled term "raptor".

Cheers!!
Papers
Brink, K.S. et. al. (2015) Developmental and evolutionary novelty in the serrated teeth of theropod dinosaurs.
Scientific Reports 5, article no. 12338, July 2015
Gignac, P. M., Makovicky, P. J., Erickson, G. M., & Walsh, R. P. (2010). A description ofDeinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations. Journal of Vertebrate Paleontology, 30(4), 1169-1177.
Kim, J.Y., Kim, K.S. and Lockley, M.G. 2008. New didactyl dinosaurs footprints (Dromaeosauripus hamanesnsi ichnogen. et ichnosp. nov.) from the Early Cretaceous Haman Formation, south coast of Korea. Palaeogeography, Paleoclimatology, Palaeoecology 262: 72-78
Li, Rihui., Lockley, M.G., Makovicky, P.J., Matsukawa, M., Norell, M.A., Harris. J.D., Liu, M., (2007) Behavioral and faunal implications of Early Cretaceous deinonychosaurian trackways from China. Naturwissenschaften (2008) 95: 185-191 online
723–730.http://www.xinglida.net/pdf/XING_et_al_2012_Deinonychosaurians_Tracks.pdf
Support me on Patreon.
"A Long habit of not thinking a thing wrong, gives it a superficial appearance of being right, and raises at first a formidable outcry in defense of custom". Thomas Paine
Support me on Patreon.
Like antediluvian salad on facebook. Visit my other blog southlandbeaver.blogspot
Watch me on Deviantart @NashD1. Subscribe to my youtube channel Duane Nash.
30 comments:
Interesting hypotheses
I actually compared dromaeosaurs to large mustelids for a long time.
Nice to see that dromaeosaurs, despite the fact some of them were clearly not as fast, definitely were not sluggards. Speed isn't mobility.
Right if those fossil trackways represent cruising speeds and they were not actively chasing something anything more than 4 mph is well above what most animals normally walk at. In one day they might cover 20-30 miles, that is getting around.
An amazing "saga" of posts to sum up such amazing organisms. I don't have much else to say/ask about them seeing as how you covered nearly everything about their general behaviour, ecology, etc.
As I've talked to you about before, the large scale Dinosaur field guide I'm working on uses common names just like modern animals. I never wanted to use the word "raptor" for Dromaeosaurs since it's already taken by another group of highly carnivorous Theropod, so what are some good ideas that you (or anyone else in the comment section) have in mind that I could use? I thought of the name "Eagledrak" (literally Eagle Dragon) as a common name, but I have also thought of the name "Hell Hawk" to either replace or to accompany Eagledrak as a common name. Got any other suggestions?
"Hell-pacer" "Running Bone Rattler" "Demon-scarf-hell-face" LOL
A couple of notes:
There *IS* a Cloverly tyrannosauroid: http://www.tandfonline.com/doi/abs/10.1080/08912963.2010.543952#.V6SD4GUxu3c
In terms of metatarsal/tibia ratio, and scaled for size, even Saurornitholestes is actually fairly stumpy-footed.
"
There *IS* a Cloverly tyrannosauroid: http://www.tandfonline.com/doi/abs/10.1080/08912963.2010.543952#.V6SD4GUxu3c"
I think the problem is more to do with:
Is the size and shape of the tooth resembles the scar made in tenontosaurus bone(Deinonychus does)? Even if it does, that means that Tyrannosaurs had a blade-tooth unlike later tyrannosaurs.
I think I may have sounded rude up there. What I was trying to say is that is there good reason to believe that Tyrannosaurs with classic blade tooth(that resembles Deinonychus tooth in size and shape) were capable of making significant bone damage via bite force.
Duane I actually really love the names Hell-pacer and Bone Rattler. I'll be using these if that's fine with you ;).
Also, are there any other indications of other Dromies besides Deinonychus using this "bone rattling" technique? I'd be suprised if other well known genera weren't doing the same.
And speaking of bone rattling, do you think another common named for various Dromie taxa could be "Bonepeckers", in the same vain as modern Woodpeckers?
@khalil any indictions of other dromies using this "bone rattling" technique? Not that I know of and it is far from proven or even adequately tested in Deinonychus. It's a hypothesis, one which may potentially explain the bone puncture marks, although I would not get too expectant in people jumping on board with it cuz skeptical scientist is skeptical.
@Thomas Holtz Point taken regarding size correction for saurornitholestes and ref on the Cloverly turannosauroid. Anonymous iterated my concerns with simply ascribing the bone damage to a relatively rare and unknown tyrannosauroid.
I did in the post emphasis the long range loping ability of the wolverine - a stumpy footed carnivoran - which never the less travels staggering distances and has immense home ranges. I did make the analogy with that mustelid because dromie "cruising" foot print speed is often times not too shabby considering their stumpy footed design and the rarity of actual chases documented via footprints. Such high cruising speeds should stir interest in the use of the caudal rods as potential elastic rebound organs in facilitating such efficient paces over long distance as hunter - scavengers of often times marginal habitat (i.e. deserts/dunefields).
Hi Duane -
If you really are interested in understanding dromaeosaurs, you should be interested in my book where I ... SOLVED THESE QUESTIONS - not just questions to do with the tail. Far from my account being the only one that seems to be completely ignored, it is in fact the only one you need to read.
Would you like me to send you a free e-copy? I think it's about time you read it.
I'd actually put the tail explanation out on the internet several years before I published the book four years ago, but of course the "Ugly Sisters" complex amongst so many, made people want to pretend that I and my comments didn't exist. Energy storage is an interesting idea, which is why I noticed it and formed it into my theory. But seeing as many flying dromaeosaurs such as Microraptor gui had an extremely long stiff tail but clearly, from their feathered feet, could not have been long-distance foot predators, and since no predatory theropods without the special claw had the tail, it's unlikely energy saving in locomotion is the answer.
But energy utilisation by the tail is instead to do with the teeth. And I also explained why the back of the teeth had the larger denticles. And I also explained how the rear wings worked. And I also explained why it was the descendants of dromaeosaurs that won out in bird survival in the end.
Palaeontology usually seems to be played as a social game which pretends to be science, but if you choose you can actually follow the real science, Duane!
"I did in the post emphasis the long range loping ability of the wolverine - a stumpy footed carnivoran - which never the less travels staggering distances and has immense home ranges."
Although, I have to mention that Wolverine seems to be a plantigrade, so being efficient cruiser is not that strange. Same goes for human and bear.
Hi Mr. jackson
Thanks for comments and my email is duanen8@gmail.com if you want to send me an e-copy
(1) "seeing as many flying dromaeosaurs such as Microraptor gui had an extremely long stiff tail but clearly, from their feathered feet, could not have been long distance foot predators
and
(2) since no predatory theropods without the special claw had the tail, it's unlikely energy saving in locomotion is the answer"
First off in point (1) I am not so sold on the notion that just because Microraptor and other microraptorines had feathered hindlimbs that this in fact hindered or negated their terrestrial capability. Feathers can be especially tough and resilient and I don't see compelling reason why hindlimb feathers were so dainty and weak that brushing past vegetation or active movement would have been discouraged.
Point (2) I need to reiterate that dromie tails share remarkable similarity to rhamphorynchine tails. Why is this? Dromies were once flighted and it seems such a tail morphology had something to do with flight... did it aide in balance,,, did it store and recoup energy from the flight stroke?... I don't know. But what I am hedging to is that the caudal rod morphology inherited from flighted ancestors was exapted into a terrestrial capability in terrestrial dromies. The reason that other theropods did not evolve a similar caudal rod tail adaptation is quite simple - they did not have flighted ancestors to inherit the basic design. What they had - and other theropods/dinosaurs could have had parallel solutions to movement efficiency - was just good enough. Not perfect - but good enough.
I will have piece on microraptorines up soon so I will say more on them in the future...
I am interested in reading your book although I might disagree with some of your ideas/conclusions. But in this way I am treating you no different than many of the "luminaries" of the field which I also differ from on many points.
Cheers
Interesting as always, Mr. Nash. I feel several comments are in order.
1. I feel that the Manning et al study has been too quickly accepted as dogma. Take a look at figure 9 in Kirkland's original description of Utahraptor: https://www.academia.edu/225747/A_large_dromaeosaurid_Theropoda_from_the_Lower_Cretaceous_of_Eastern_Utah
You'll notice that Utahraptor's killing claw is teardrop-shaped in cross section, and that it retains this shape very nearly all the way to the base. It practically screams "I had a sharp edge in life!" I'm not sure whether or not this is the case with Deinonychus and Velociraptor as well, but if it is, Manning et al's justification for testing a claw with a dull ventral surface on the basis of phylogenetic bracketing seems extremely weak - especially so given that similar arguments could be constructed demonstrating that dromaeosaurs probably didn't have those ossified ligaments in their tails, or that they probably didn't have serrated teeth! Why no one in the scientific community has pointed out this seemingly obvious shortcoming of Manning's test is beyond me. Suffice it to say that the slashing hypothesis is far from falsified. This isn't to say that the RPR model is totally wrong. After all, an entire clade of today's apex predators use ventral keeled claws for prey restraint. Whether slashing or prey capture was the primary use of the claw remains to be determined.
2. Re. Your "tooth boring" hypothesis:
a) given the orientation of the tooth in the mouth, it looks to me that it would be the anterior serrations that would be doing most of the work, not the posterior ones.
b) I'm somewhat confused as to the benefits of such a feeding style. The potential benefits of the bone saw shimmy you proposed for carnosaurs were fairly obvious. What purpose would this "bone-drill dougie" serve?
c) it seems to me that there's a simpler explanation for how a Deinonychus could have made such marks. Energy is force times distance, therefore force is energy divided by distance. Energy is conserved, so if you apply a force of 1000 Newtons to something (for example, a mandible full of teeth) for 20 cm, and then something else (for example, a dinosaur bone) decelerates the first thing for a distance of 5 cm, that something else will experience a force of some 4000 Newtons.
It's probably worth mentioning that, in a scrum of feeding dromaeosaurs, a lot of animals would be jostling to snap up a chunk of meat. It's probably also worth mentioning that some of these quick, hard, snapping bites might be poorly aimed, and could thus end up gouging marks in bones far deeper than would be expected from the sustainable force output of a dromaeosaur's jaws.
For whatever it's worth, to my (admittedly amateur) eyes, this explanation of the tooth marks seems to fit in better with the whole "dromaeosaurs as combative scavengers" model than your "bone-drill dougie" hypothesis does.
3. Regarding the use of the term "raptor," seeing as falcons, owls, accipitridae, and new-world vultures form a polyphyletic group, I see no reason why extinct clades of predatory/carnivorous bird - including at least dromaeosaurs and the terror birds - should not be subsumed under the term as well.
†hanks for comment Dave
1) Interesting you mentioned a sharp edge likely in Utahraptor as I have came across this notion elsewhere in addition to new thought on the subject... well my thoughts have been evolving on the killing claw and I may have to eat some of my own words and come back around to the killing claw actually being a killing claw.
2) a) "looks to me the anterior serrations that would do most of the work" I think that they both had a function. The diminished anterior serrations allow piercing while the posterior serrations which are apically hooked do the excavation. I'm sure a computer simulation could be used to test this.
b) bone drilling would be useful not just for bone but more importantly getting into the thick hides of dinosaurs. Once the teeth get "drilled" into the carcass then the neck and body could pull back with better purchase. The "bone drilling" and "bonesaw" hypotheses might be better understood as ways to penetrate and get through the tough, often times osteoderm studded hides of dinosaurs. Of course "skinsaw" or "skin- drill" don't sound as cool ergo I use the bone terminology. Dinosaur skin and hide was a daunting task to get through and these speed & friction based biting methods provide hypotheses on how best to get through the skin of animals very similar to komodo dragon or crocodile skin.
c) yes but we still have the question of relatively weak bite force in Deinonychus - where is the energy coming from? If it is not coming from static jaw pressure maybe the body is generating the energy which gets transmitted through the teeth?
3. Little known fact: Jurassic Park made the term "raptor' popular because they followed the taxonomy of Gregory S. Paul. GSP sunk Deinonychus into the genus of Velociraptor an idea that pretty much no one follows except for Jurassic Park. JP really wanted more of a Deinonychus style villain - not a coyote sized Velociraptor - in their movie. They took Deinonychus (which they assumed as a type of Velociraptor per GSP taxonomy) and made it bigger, Incidentally Utahraptor discovered later that year!! The term "raptor" therefore came about and was popularized for a mistake!!
Thank you for taking the time to respond my comments, mr. Nash.
1. Not that I think you give a crap about my opinion, but it's nice to see a willingness to admit mistakes combined with a willingness to come up with out of the box hypotheses - especially when it comes to paleo-ecology, where we have such a dearth of empirical data.
2. I'm not going to continue with the further subdivision of this heading, as I wish to transition to a holistic discussion of tooth mechanics. In deinonychus, the teeth point backwards, and the denticles point towards the tip - in other words, backwards as well. If most theropods had teeth for slashing and sawing, dromaeosaurs had teeth for gripping and pulling. Of course, the anterior serrations mean that sawing is still viable, but their reduction could indicate that it was somewhat less important than the backwards yank. As for where the energy was coming from - it was coming from the weak jaws!
Deinonychus had a 40 cm skull. Conservatively, it could have had a gape of some 35 cm. Therrian et al calculated its sustainable bite force to be 1450 Newtons. That probably couldn't be sustained through the jaw's entire range of motion, as the jaw muscles would be further from perpendicular to the tooth row at wider gapes, so we'll go with a figure of 750 N. We'll subtract 5 cm from the gape to fit in the tenontosaur bone, and cut out an additional 2.5 cm to leave room for deceleration. So our 750 N will be applied for 27.5 cm. That means that the tooth row will have an energy of 206.25 J. If the bone decelerates the tooth row for a distance of 2.5 cm, it will be experiencing a force of some 8250 N - well in line with the experimental data. Needless to say, if the muscles of the neck and legs are recruited as well - say, if the animal violently thrusts its head into the scrum to snap a bite out of the carcass - the figure could be further inflated. In my opinion, this sort of suggestion has two advantages over yours: first, in my admittedly inexpert estimation, it seems less biomechanically questionable. Second, it fits in with your whole "combative scavenger" model somewhat more organically.
I hope that my contributions are helping you think through your ideas!
Well no I do give a crap about other's opinions, in fact that was one of my main imperatives with this blog. Suggest new ideas, put them out there, and see how people respond to them either through valid critiques or other lines of evidence that I didn't think of. Sort of a group sourcing of paleo thought. A lot of my ideas of course need more rigorous and quantitative testing (math not my strong suit) and hopefully paleontologists with better experimental technology (FEA, actual models, computer simulations) will take up some of the hypotheses I have been suggesting.
We should not throw out unpolished gems just because they need a little work!!
Uggh it's been a long time since I did any physics but if your math holds up the scenario you put forth is much more reasonable than what I suggested. Which begs the question why didn't the pros think it through like you did? Admittedly using power and speed garnered from the legs and necks seems more reasonable from a biomechanical perspective but also from an evolutionary perspective - a problem with my idea is that I don't have an evolutionary mechanism - an exaptation - that would facilitate such a method as I proposed.
Don't worry I am not very wed to the idea of bone drilling teeth in dromies just kinda throwing shit at the wall to see what sticks!!
Thanks for your input Dave like I mentioned in the last post I did come across some rather novel and interesting thought on the dromie killing claw that infers a strong cutting edge and a rather bizarre use of the tail. Problem is that this idea comes from a man who is.... well he has a strong allegiance to an idea that is not very well liked in the dino-bird-paleocommunity. Unfortunately I fear this unyielding insistence on a particular idea has caused people to overlook - or not even read - his rather good idea on dromie killing claws. It's a tough needle for me to thread.
One weakness of my suggestion would be that such forces could only be delivered for a fraction of a second.if sustained pressure were required to produce such markings, my idea wouldn't work.
Would you be willing to direct me to this new thought on the killing claw?
I found this old blog post basically laying it all out there: http://lefthandedcyclist.blogspot.com/2012/02/raptors-revisited.html
Interestingly this idea presented on the blog converges with pretty much the same idea developed by notorious Birds Came First advocate John Jackson in his self-published book The Secret Dino-Bird Story.
I don't agree with the BCF idea, nor with several of Jackson's other ideas. But I do suspect he is on to something with the hypothesis he developed. Unfortunately due to his reputation online and the lack of support and evidence for BCF I don't think many are aware of his ideas on the killing claw, However when his idea is decoupled from BCF no reason for it to not attract attention and further analysis.
I will probably blog on this "pierced from within" hypothesis eventually.
Very interesting... The dromaeosaur tail is such a curious adaptation.
Out of curiosity, does the Gignac study on Deinonychus bite force happen to mention how long force has to be applied in order to produce the observed indentations? In the interest of Scientific Progress (tm), I'm interested in finding out whether my suggestion really is plausible.
to tell you the truth I don't know the ins and outs of the Gignac study just that it is referred to a lot in reference to bite force in Deinonychus.
When I read in these dromies post about the heel pads that may impair grasping ability in the feet, as well as the diminished second toe claw in more cursorial species, I started thinking about cassowaries. They also have that enlarged talon on the second toe, and they can jump up to about 5 feet in the air. They don't have a need to grasp anything since they're ground-dwellers and mainly frugivorous, but they do charge and kick out at anything they feel threatened by, capable of leaving huge gashes in the offender/victim (I've heard of one report of a cassowary ripping down a car door and leaving it with a 6-inch gash). I was thinking that perhaps dromies (perhaps the cursorial species in particular) employed this tactic in prey capture along with, and possibly preceding, biting and wing-pummeling. Thoughts?
Hi Lam Luoung thanks for comments. I tend to think that the cassowary/dromie comparison is taken too literally. The morphology of their claws are indeed very different. Indeed my thoughts on how the killing claw functioned has changed quite dramatically as of late, deviating from ideas I have expressed on these posts, as I have come across some alternative ideas on how the claw may have functioned. Stay tuned.
Again, fantastic work. Now, I must contend with your premise that the vast majority of dromies were strictly cursorial. Whichever came first, the "chicken" or the dromie - in regards to secondary flightlessness - it goes without saying their close relationship to aves. Therefore to infer that at least the first dromies were arboreal in nature would make perfect sense, as tree climbing, dwelling, and perching would be a step towards gliding and then flight. In the BCF scenario, dromies would have likely retaind enough of their arboreal adaptations to reverse the trend. Dienonychus especially, with it's highly curved claws and strong grasp, would have been highly adapted to an arboreal lifestyle. Weren't studies done to the musculature of the legs that determined this already? I need the reference! I will say I think Ostrom was right about something, as their boney, stiff tails were a perfect balance beam, maybe not for standing on one leg and karate-kicking, but for perching. I would not get too hung up on the "dienonychus" tooth bite, as I'm sure the consensus in paleo-circles is that it was at most a sign of scavenging behavior. I find it highly unlikely to think dienonychus ever hunted tenontosaurs. I know people go back and forth about their bite force, about the only way one could kill such prey would be with either a strong bite or something like your vibration theory, but only when applied to (1) the neck as in felids, or (2) the underbelly. You could make an argument where dienonychus was adept to pouncing on it's prey, attaching itself with both it's pedal and manual unguals, and attacking these two regions with their jaws. However, that is a highly specialized tactic and I doubt it was the case. Their teeth looked like they could handle moderate prey, but not necessarily that big. I also heard a theory worth mentioning where dromies such as these fed off prey in this manner only parasitically, they did not intend to kill it's prey but instead partially eat some of it's soft tissue, allow it to live only to feed another day. Maybe. But back to my core thoughts, I believe the term eudromaeosaurid should apply only to dromaeosaurinae and exclude velociraptorinae. Those were the true hunters. There is no doubt they could have been efficient hunters and the main killing weapon were those incredibly powerful, tyronnosaurid-like jaws. Look at those teeth. But what about those pesky claws no one shuts up about. If it is a hold-over gene from climbing trees, why is it expressed in these cursorial dromies? Because yes they always played a pivital role in prey capture (RPR) as well. However, these claws are blade-like and better suited for hunting, and hunting only. For years I thought they had to be of a theriznosaur or perhaps even allosaurid. I did not think that thing could exist, I really was shocked to hear it wasn't a chimera. Imagine a utahraptor in this manner. It's jaws are so massive with the lower being curved its as if it could chew its way into it's preys body cavity. Being firmly attached with it's claws, its robust size would exhaust prey immediatly. They look like they would just be dieing to scap with whatever. Either way, it's jaws we're much more efficient for dispatching prey than pummeling them with it's manuals. Yes dromies are the longest tenured branch of terrestrial hunter/scavenger we have ever seen. It was their ability to adapt. They took all different shapes, sizes, and niches. They never took one form and ran with it. Speak more to the diversity in the group or you have to be more precise as to if you are referring to just eudromies or all dromies. Thanks so much.
If i had any balls i would be willing to bet money that this whole utahraptor thing is a farce. Something seems amiss about the utahraptor project. Among the initial controversy that it wasnt even a Dromaeosaurid, Kirkland kept pointing out "known Dromaeosaurid characteristics" of the skull. Im looking at the same skull, i just dont see it. That tells me kirkland is dancing on a very thin tight rope trying to connect those bones to the Dromaeosauridae . Why? Because there is financial incentive to fawn the flames of the idea of giant Jurassic park killing raptors (i.E. Bakkers book " raptor red). The public gobbles it up. Look at the updated skelatel. Other than the arrangement of the pedal and manual unguals-which could be misinterpretated-it does not look like a dromie at all. No boney rods in the tail. Leg bones more than 2X's as thick as allosaurus. My guess is the utahraptor project is kirklands last chance to cash in before this animal is exposed for what it is. If so, the whole conception of the Dromaeosauridae comes crashing down . Hope im wrong.
@Anonymous Yeah I'd have to push back on this notion a lot. I've met Kirkland he has done a lot of great work on American public lands and for dinosaurs. He doesn't strike me as the showboating type or in it for the money type you are suggesting. For example he is more than accommodating towards his grad students and their presentations at the SVP and doesn't try and overshadow or overstate his involvement in their work. In fact paleontologist generally don't get into this for the money. I don't agree with some of his ideas about dinosaurs BUT I do think he is an outstanding geologist, field worker, and overall a class act based on what I have seen and heard. Something tells me he wouldn't want to risk his reputation and life's work for misrepresenting what Utahraptor actually is.
More to the point I don't think loads of money is pouring into the project from a public insatiable for dromaeosaurids.
Regarding the different look to this animal compared to other dromies I think we should expect such diversity within such a long tenured family.
Best,
Duane
Well ive never seen paleontologists call for public donations quite like this. Im not saying he is nefarious in any way, but he does strike me as a sensationalist.
Lol, i realize the public does not have an "insatiable" thirst for dromaeosaurids, or the "raptors" from jp, but a very small minority does. My point was many paleontologists, not just kirkland, have likened dromies to the jp raptors to help garner public interest. For decades, they have been depicted as pack hunting, claw slashing killing machines when there was literally no supporting evidence to back it up. And yes, i do believe there is more of a financial incentive (such as books, documenteries, etc) to "discover the real raptors from JP"-which is exactly what utahraptor was originally touted to be-as opposed to a primative tyrannosaurid. Do you really think there would even be a utahraptor project at all if it belonged to another branch of theropod? I will close with one point we all will agree on, that it seems as more time goes by, the less we know about these creatures. We are just as much in the dark on this topic today as we were in 1964.
Post a Comment